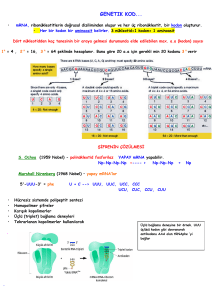
Translasyon
Doç. Dr. Nurten Özsoy
mRNA tarafından taşınan bilgilerin protein dizilerine aktarılmasıdır
sitoplazma
serbest amino asitler
Gen protein zinciri
Sentezlenen
mRNA
tRNA
ribozom
Transkripsiyon
DNA
3
RNA
polimeraz
RNA
transcript
5
Ekson
RNA’nın işlenmesi
RNA transcript
(pre-mRNA)
Intron
Aminoaçil-tRNA
sentetaz
NUKLEUS
Amino
acid
Başlama kompleksinin
oluşumu
Sitoplazma
mRNA
AMİNO ASİD AKTİVASYONU
tRNA
Büyüyen
polipeptid
Aktive edilmiş
amino asid
A
P
E
Ribozom
altbirimi
5
TRANSLASYON
A
E
Kodon
Ribozom
Antikodon
3
Genetik kod: mRNA üzerinde 5'3' yönünde nukleotidlerin sıralanması
Kodon – mRNA da, 5'3' yönündeki üçlü nukleotid dizileri
Antikodon – tRNA’larda, mRNA’lardaki kodonlara baz eşleşmesine
uyacak şekilde, üçlü nukleotid dizileri
DNA
Gen 1
Gen 2
Gen 3
Amino asid
antikodon
kodon
Genetik kodun özellikleri
Genetik kod evrenseldir (tüm canlılarda aynı kodonlar kullanılır).
Genetik kod 4 farklı bazın üçlü dizisi şeklinde olduğundan 43=64
kodonu içerir.
Üçlü kodonun son nukleotidi değişkendir (wobble).
mRNA (3') 3 2 1 (5') (kodon)
(diziler 5' 3' okunur)
tRNA (5') 1 2 3 (3') (antikodon)
Kodonun üçüncü bazı antikodonun ilk bazına uyar.
Antikodonun ilk bazı
Kodonun 3. (wobble) bazı
tRNA 1
1
2
2
3 kodonu tanıyabilir
Yirmi amino asitten iki tanesi; metiyonin ve triptofan birer kodona sahiptir.
Geri kalan amino asitler için iki, üç, dört veya altı kodon mevcuttur.
Bir amino asidin birden fazla kodona sahip olması durumuna dejenere olma
denir.
Genetik kodun dejenerasyonu
Amino asit
Kodon sayısı
GENETİK KOD
Örneğin antikodonu (5') ICG olan tRNAarg üç farklı arginin kodonu
tanıyabilir
3 2 1
3 2 1
tRNA (antikodon)
(3') G-C-I
G-C-I
mRNA (kodon)
(5') C-G-A C-G-U
1
2 3
1
2 3
3 2 1
G-C-I (5')
C-G-C (3')
1 2 3
Francis Crick’in wobble baz hipotezine göre:
1. Kodonun ilk iki bazı antikodondaki bunlara uyan iki bazı ile WatsonCrick baz çifti oluştururlar ve amino asitlerin spesifisitesini tayin
ederler
2. Antikodonun 1. bazı, tRNA tarafından okunan kodon sayısını belirler.
Antikodonun
1.
bazı
C
U
I
veya
veya
A
G
-
tRNA bir kodonu
tRNA iki kodonu
tRNA üç kodonu tanır.
3. Üç kodon bir amino asidi belirlediği zaman sadece ilk iki bazı farklı
olan kodonların tRNA’ları farklıdır.
4. Amino asidler için 3 kodon şifreleme yapmaz - UAA, UAG, UGA.
Bunlara anlamsız kodonlar denir. Hücrede en az 2 tanesi sonlandırma
sinyalleri olarak kullanılır. Geri kalan 61 kodon amino asidleri kodlar.
6. Genetik kod üst üste gelmeyen bir özelliktedir. Genetik kod’ta
kodonları ayıran nokta, virgül gibi kesinti yoktur ve mesaj anlamsız bir
kodona erişene kadar üçlü bazlardan ibaret bir dizi şeklinde okunur.
Kodonlar mRNA üzerinde birbirini izleyerek yan yana dizilmiştir.
Sıranın bir nukleotidinin kayması bile önemli protein değişikliklerine
neden olur.
7. AUG hem başlangıç
metiyoninlerinin kodonudur.
metiyonin’in
kodonu
hem
de
ara
8. 61 kodon 20 amino asid için eşit ve dengeli bir şekilde dağılmamıştır.
mRNA’daki herbir amino aside özgü olmak üzere 2, 3 veya 4 gibi daha
çok sayıda farklı üçlü diziler saptanmıştır.
Kodonların ilk 2 bazının bir aile oluşturduğu düşünülürse, bu 64
muhtemel kodonun 16 aile oluşturabileceği anlaşılır. Bazı ailelerde 4
kodon ayni amino asidi şifreler. Bunlara karışmamış aileler denir. 16
kodon ailesinin 8’i karışmamıştır. Bir amino asidden fazlasını şifreleyen
kodon aileleri ise karışıktır (6 aile). Geri kalan 2 aile de UG ve AU
aileleridir.
Ribozomlar
Ökaryotik
Prokaryotik
Proteinler
Alt birimler
Ribozomlar
Protein biosentezinin mekanizması
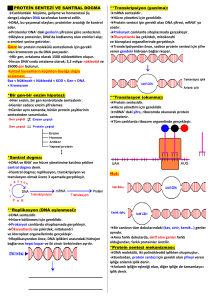
1. Amino asidlerin aktivasyonu (sitoplazmada gerçekleşir)
Amino asid + tRNA + ATP Aminoaçil-tRNA + AMP + PPi
Amino asid
adenozin
Her amino asid için özel bir enzim bu amino asidi
ATP
aktive eder. Aktivasyon için gerekli olan kimyasal enerji
adenozin
pirofosfat
Pi
fosfat
.
Aminoaçil-tRNA sentetaz
EnzimaminoaçilAMP
Pi
tRNA
adenozin
AMP
Aminoaçil-tRNA
(aktive edilmiş amino asid)
ATP’tan sağlanır. Bu esnada enzimle birleşmiş olan
amino asidin karboksil grubunun ATP ile reaksiyona
girmesi sonucunda enzim-aminoaçilAMP meydana
Aminoaçil gelir ve PPi açığa çıkar. Bu suretle meydana gelmiş
tRNA
aktive edilmiş amino asidi yine aynı enzimler tarafından
tRNA ile birleşerek aminoaçil tRNA oluşur.
E.coli
Aminoaçil-tRNA
sentetaz
Aminoaçil -tRNA
Adenin
Enzim- aminoaçilAMP (karışık bir asit anhidrit’tir)
Adenin
Sınıf I sentetazlar
(monomerik)
RNA’nın
3' ucu
Sınıf II sentetazlar
(dimerik)
Adenin
Adenin
Adenin
Adenin
Adenin
Adenin
Transesterifikasyon
2'-O -aminoaçil tRNA
3'-O-aminoaçil tRNA
tRNA’nın 3' ucu
Aminoaçil grubu
Amino asit
kolu
3'-O aminoaçil tRNA
kolu
kolu
Antikodon
kolu
Enzim (aminoaçil-tRNA sentetaz) + Amino asid + ATP Enzim-aminoaçil AMP + PPi
Enzim-aminoaçil AMP + tRNA Aminoaçil-tRNA + Enzim + AMP
Enzim + Amino asid + ATP + tRNA Aminoaçil-tRNA + Enzim + AMP + PPi
Başlama
• Başlama kodonu AUG’dir
• Her protein sentezi metiyonin ile başlamaktadır.
• Hücrede metiyonin için özel iki tRNA türü bulunmaktadır.
- protein molekülünün içinde herhangi bir noktada yer alan metiyonini
taşıyan tRNA – Met-tRNAMet şeklinde gösterilmektedir.
-Met-tRNAfMet (formilmetiyonil-tRNA) şeklinde gösterilen tRNA
molekülü, mRNA üzerindeki doğru başlama bölgesini tanıyabilmektedir
(başlama kodonuna bağlanmaktadır)
Metiyonin + tRNAMet (fMet) + ATP
Metiyonil-tRNAMet (fMet) +AMP + PPi
Her ikisinin aktivasyonunu aynı metiyonilaçil-tRNA sentetaz gerçekleştirir
Transformilaz
Metiyonil-tRNAfMet +AMP + PPi
N10-Formil THF
N-Formilmetiyonil-tRNAfMet
THF
(fMet-tRNAfMet)
Prokaryotlarda metiyoninin fMet-tRNAfMet molekülüne bağlandıktan
sonra formillenmesinin başlıca iki amacı bulunur.
1. Sentez başlarken ribozomun peptidil (P) bölgesinde bulunabilen tek
aminoaçil-tRNA olmasını sağlamak.
2. fMet-tRNAfMet ‘nin mRNA içinde bulunan metiyoninin kodonları için
kullanılmasını engellemektir.
N-formilmetiyonin
Prokaryotlar
Polipeptid zinciri
Ribozom
Aminoaçil
-tRNA
Prokaryotlarda sentezin başlaması
Sentez polipeptid zincirinin N-ucundan başlar
1. 30S ribozom altbirimi
2. Sentezlenecek polipeptidi şifreleyen mRNA
3. Başlangıç fMet-tRNAfMet
4. Başlama faktörleri (IF-1, IF-2, IF-3)
5. GTP
6. 50S ribozomal altbirimi
7. Mg2+’ye gereksinim vardır
Başlama basamağı 1
mRNA
Üç bölgeli ribozom modeli (Danesh Moazed ve Harry Noller
tarafından önerilmiş)
P bölgesi – uzayan peptid zincirini
A bölgesi – gelen aminoaçil-tRNA molekülünü taşır
E bölgesi – aminoaçil grubunu veren tRNA molekülünün
ribozomu terketmeden önce beklediği bölgedir.
mRNA molekülünün 5' ucuna küçük ribozom alt birimi molekül boyunca
ilerleyerek, fMet-tRNAfMet molekülünün antikodonu ile baz eşleşmesi
yapabilen AUG kodonuna geldiği zaman protein sentezinin başlama
noktasını bulmaktadır.
Bir ribozomda dört tane RNA bağlanma bölgesi vardır: Bunlardan bir tanesi mRNA için, üç
tanesi de (A, P ve E) tRNA içindir.
Oluşan mRNA’nın AUG şifresinden ribozoma bağlandığı konusunda J. Shine ve L. Dalgarno
tarafından 1974 yılında bakteriler üzerinde yapılan bir çalışma ile tespit edilmiştir. Bu çalışmanın
sonucu olarak mRNA’nın 5'-uç kısmında yer alan başlatıcı kodon AUG’den yaklaşık 10 nukleotid
öncesinde AGGAGGU bazlarından oluşan ve Shine-Dalgarno dizisi olarak isimlendirilen
bölgenin ribozomun küçük alt birimindeki 16S rRNA’nın 3' uç kısmında uyumlu bir bölgeyi
(UCCUCCA) tanıyıp bağlandığı anlaşılmıştır.
Ökaryotlarda ise Shine-Dalgarno dizisi bulunmaz ve bunun yerine mRNA’nın 5' uç bölgesinin ilk
bazları olarak metillenmiş guanin ile oluştuğu düşünülmektedir.
Shine-Dalgarno dizisi
Sadece prokaryotlarda bulunmuştur. 4 ile 9 arasında purin kalıntısı içeren, başlama kodonunun
5' ucundan 8-13 baz çifti uzunlukta bir başlangıç sinyalidir.
Başlama basamağı 2
Büyük ribozom alt birimi küçük ribozom alt
birimine, başlama kodonu (AUG) P konumunda
olacak şekilde bağlanır
Başlama basamağı 3
fMet-tRNAfMet’nin antikodonu AUG başlama
kodonuyla eşleşecek şekildedir ve UAC’dir.
Gerekli enerji GTP’den sağlanır.
30S ribozomal altbirimi
Başlama faktörleri
GTP
30S başlama kompleksi
Basamak
1.
30S
ribozom
altbirimine iki başlama faktörü, IF-1
ve IF-3, bağlanır. IF-3, 30S ve 50S
altbirimlerinin erken birleşmesini
engeller. Başlangıçtaki (5') AUG
doğru yerine mRNA’daki ShineDalgarno dizisi ile yönlendirilir.
IF-1’in önceden A bölgesine
bağlanması, fMet-tRNAfMet ‘nin P
bölgesine yönlenmesini mümkün
kılar.
Basamak 2. 30S altbirimi, IF-3 ve
mRNA’dan oluşan komplekse hem
GTP-bağlı IF-2 ve hem de başlatıcı
fMet-tRNAfMet katılır. tRNA’nın
antikodonu mRNA’nın başlama
kodonuyla doğru baz eşleşimi yapar.
30S başlama kompleksi
70S başlama kompleksi
Basamak 3. Bu büyük
kompleks
50S
ribozom
altbirimiyle birleşirken, aynı
anda IF-2’ye bağlı GTP, GDP
ve
Pi’a
parçalanarak
kompleksten ayrılır.
Basamakların tamamlanması
başlatma kompleksi adı verilen,
işlevsel bir 70S ribozomu
oluşturur.
Uzama
• Başlangıç kompleksi (70S ribozomu)
• Aminoaçil-tRNA
• Uzama faktörleri (EF-Tu, EF-Ts ve EF-G)
• GTP gereklidir
Zincir uzaması 1
UAC
UGG
Uzama basamağı başlama kodonuna komşu,
ribozomun A bölgesindeki kodona ait amino asidi
taşıyan tRNA molekülünün aminoaçil-tRNA
şeklinde ribozoma bağlanması ile başlamaktadır.
Zincir uzaması 2
İlk aminoaçil-tRNA nın aminoaçil kalıntısı,
ribozoma ikinci olarak gelen aminoaçil-tRNA nın
amino grubuna peptidil transferaz enzimin
yardımıyla peptid bağı ile bağlanır. Bu tepkimeye
transpeptidasyon adı verilir.
Meydana
gelen
dipeptid:
Met-Thr
Peptid bağı oluşumu
Transpeptidasyon,
aminoaçil-tRNA
molekülünün
amino
grubunun,
peptidil-tRNA molekülünün ucundaki
karboksil grubu ile nukleofilik olarak
etkileşmesini kapsamaktadır. Böylece P
bölgesinde bulunan peptid, peptidil
transferaz enzimin etkisiyle A
bölgesindeki aminoaçil-tRNA ile ester
bağı (peptid) yaparak bağlanmakta ve P
bölgesinde peptidi taşıyan tRNA ise
serbest kalmaktadır.
Zincir uzaması 3
Translokasyon (yer değiştirme): Ribozom mRNA üzerinde 3' yönünde
ve translokaz enzimin etkisiyle P konumu bir sonraki kodonu
kapsayacak şekilde hareket eder ve böylece dipeptidil-tRNA A
konumundan P konumuna kayar ve deaçil-tRNA E konumuna geçer.
Translokasyon olayı için gereken enerji
translokaza bağlı GTP’ın hidrolizi ile
meydana gelir.
UAC
UGG
Polipeptid uzamasının temel döngüsü translasyonun etkin ve doğru olmasını sağlar. Her döngü
sürecinde ribozoma iki uzatma faktörü (EF-Tu ve EF-G) girer ve çıkar. Bunların ikisi de GTP’yi
GDP’ye hidroliz eder ve bu işlem sırasında şekil değişikliğine uğrarlar.
EF-Tu translasyonun hızlı bir şekilde ilerlemesine yardımcı olmanın ötesinde, yüklü tRNA ile kodon
arasındaki ilk etkileşimi de izleyerek, translasyonun doğruluğunu arttırdığı düşünülmektedir.
•İlk bağlanma işleminde, EF-Tu’ya sıkıca bağlanmış olan bir aminoaçil tRNA molekülü küçük alt
birimin A yerindeki kodon ile geçici olarak eşleşir (uzama faktörleri kodon antikodon eşleşmesine
izin verirken, amino asidin uzayan polipeptid zincirine bağlanmasını engeller).
• İlk kodonun tanınması, EF-Tu’nun üzerindeki GTP’nin hidrolizlenmesini tetikler (bu süreç yanlış
yerleşmiş olan tRNA’nın ribozomdan çıkmasına olanak verir).
• EF-Tu tRNA’yı bırakarak ribozomdam ayrılır ve protein sentezinin devam etmesine olanak sağlar
(bu süreç yanlış bir tRNA molekülünün varlığında tam eşleşmeyen kodon-antikodon çiftin bu süre
içinde kendiliğinden ayrılmasına olanak verir).
• Yeni bir aminoaçil-tRNA’nın A konumuna girmesi için EF-Tu ya bağlı olan GDP’in GTP ile
değişmesi gereklidir. Bunu sağlamak için Ts faktörü devreye girer.
Translokasyon
EF-Ts’nin
yardımıyla
EF-Tu’ya
bağlı olan
GDP’in G
ile değişm
Peptidil
transferaz
Peptid bağı oluşumu
Ribozomun mRNA üzerindeki hareketini EF-G proteini (Translok
bir mol GTP hidroliz ederek sağlar)
Zincir uzaması 4
E bölgeden tRNA hidrolitik olarak
ayrılarak sitozole geçer.
Zincir uzama basamakları tekrarlanır 1-4
Peptidin uzama süreci bu şekilde devam
eder ve olay ribozomun mRNA’nın 3'
ucundaki sonlanma kodonuna kadar
sürer.
UGG GUG
Zincir uzaması basamağı deaçil-tRNA’nın
E bölgeden ayrılmasıyla son bulmaktadır.
Sonlama basamağı 1.
Polipeptid zinciri P bölgesinde,
sonlanma kodonu (UAA, UAG
veya UGA) A bölgesindedir.
Sonlama basamağı 2.
Sonlanma veya salıverici faktörler
(proteinler) A konumundaki
sonlanma kodonuna bağlanır.
Sonlama basamağı 3.
Sonlanma faktörlerin esteraz
aktiviteleri ile P konumunda bulunan
polipeptidil-tRNA’nın tRNA-3'-O-OC….. ester bağını hidroliz eder
Met-Thr-His-Asp-Gly
Sonlama basamağı 4.
Ayrılan polipeptid ribozom
sisteminden dışarı çıkar ve 70s
ribozomu yeni bir protein sentezini
başlatmak üzere 30s ve 50s alt
birimlerine ayrılır.
Üç sonlanma faktörü (RF) devreye girer:
RF-1, UAG ve UAA kodonlarını
RF-2, UGA ve UAA kodonlarını tanır.
RF-3, iki sonlanma faktörünün bağlanmasını
GTP hidrolizi ile kolaylaştırır.
PeptidiltRNA
hidrolizi
Ökaryotlar
Primer
transkriptin
işlenmesi
Primer
transkript
Başlama faktörleri
Ökaryotlarda sentezin başlaması
Ökaryotlarda sentezin başlaması prokaryotlara göre
daha karmaşıktır. Başlangıç kompleksinin oluşması
için çok sayıda ökaryot başlama faktörü (eIF)
devreye girer.
Küçük ribozom altbirimi, 5' kep yapısını ve
40S altbirimi +
başlama faktörleri
üzerine bağlı olan eIF-4E, eIF-4G ve eIF-4A
faktörlerinden oluşan eIF4F’i tanıyarak mRNA
molekülünün 5' ucuna bağlanır. Bundan sonra,
küçük altbirim mRNA boyunca ilerleyerek (5'
60S altbirimi
Başlama
faktörleri
den 3' ne doğru) ilk AUG’yi arar. Bu işlem
ATP’den aldığı enerji ile küçük altbirimin RNA
ikincil yapısı boyunca tarama yapmasına olanak
veren helikazlar gibi ilave başlangıç faktörleri
(eIF-4A) tarafından desteklenir. AUG kodonuna
geldiği
80S başlama kompleksi
zaman
başlama
altbiriminden ayrılır.
faktörleri
küçük
Ökaryot mRNA’larının 3' ve 5' uçları,
çeşitli başlama faktörlerini ve poli(A)’ya
bağlanan protein (PAB) içeren protein
kompleksiyle bağlıdır.
Başlıklı ucu tanıyan ve oraya bağlanan eIF4E, eIF-4G ve eIF-4A faktörlerinden oluşan
eIF4F’dir.
eIF-4G, hem eIF-4E hem de PAB’ye
bağlanarak bunları bir arada tutar. RNA
helikaz aktivitesine sahip olan eIF-4A
yardımı ile mRNA’nın ikincil yapısı
açılarak, ribozom altbirimlerine ulaşmak
mümkün olur.
Gen
Tercüme edilmemiş
bölge
Ökaryotlarda zincir uzaması
Uzama faktörleri: eEF-1 ve eEF-2
eEF-1 – aminoaçil-tRNA molekülünü ribozomun A bölgesine
yerleştirir
eEF-1 – GDP ile GTP’nin yer değiştirmesini sağlayarak eEF1’nın yeniden döngüye katılmasına yardımcı olur.
eEF-2 – GTP hidrolizi ile birlikte translokasyonun
gerçekleşmesini sağlar.
Ökaryotlarda zincir sonlanması
Ökaryotlarda tüm bitiş kodonlarını tanıyan tek bir sonlanma
faktörü (eRF1) bulunur. eRF1, bir GTPaz olan eRF3 ile birlikte
sonlanmayı gerçekleştirir.
Protein sentezinin bilançosu
Amino asit aktivasyonu
- 2ATP
(ATP AMP + PPi
PPi Pi + Pi)
Aminoaçil-tRNA’nın A konumuna girişi
- 1ATP
(GTP GDP + Pi)
A konumundaki yeni oluşmuş
peptidil-tRNA’nın P konumuna
translokasyonu
- 1ATP
(GTP GDP + Pi)
Her peptid bağın oluşması için 4 yüksek enerjili bağın yıkılması gerekir.
Prokaryot ve ökaryotlar arasındaki farklılıklar
Bakteri
• Ribozom: 30S+50S →70S
• Başlama (initiation) faktörleri
– IF1, IF2, IF3
• Uzama (elongation) faktörleri
– EF-Tu, EF-Ts, EF-G
• Sonlanma (release) faktörleri
– RF1, RF2, RF3
• mRNA’ nın 5' - başlığı yok
• 30S ribozom altbirimi ShineDalgarno dizisine (5'-AGGAGGU-3')
bağlanır
• Translasyon transkripsiyonla
eşzamanlı olarak gerçekleşir
Ökaryotlar
• Ribozom: 40S+60S → 80S
• Birçok başlama (initiation)
faktörleri (14 faktör)
– eIF1, eIF1A, eIF2, eIF2B, eIF3,
eIF4A, eIF4B, eIF4E, eIF4F,
eIF4G, eIF4H, eIF5, eIF5B,
eIF6
• Uzama (elongation) faktörleri
– eEF1, eEF1, eEF2
• Sonlanma faktörleri
– eRF1 (veya eRF2), eRF3
• Ökaryot mRNA’ların 5' ucunda
başlık, 3' ucunda poly(A) kuyruğu
bulunmaktadır
• 40S altbirimi 5'- başlığına veya
poly(A) kuyruğuna bağlanmaktadır
• Translasyon sitoplazmada meydana
gelmektedir
RNA polimeraz
DNA
mRNA
Poliribozom
Hem ökaryotik, hem de prokaryotik hücrelerde protein sentezinde
10 ile 100 ribozom aynı anda aktiftir. Bunlara polizom denir.
Transkripsiyon yönü
RNA polimeraz
DNA
Poliribozom
Polipeptid
(amino ucu)
Polizom
Ribozom
mRNA (5' ucu)
Protein sentezinin antibiyotikler ile denetimi
I’m complicated but once
you get to know me
I’m really pretty nice.
Any questions?
5’
3’