Gen İfadesi - mustafaaltinisik.org.uk
advertisement

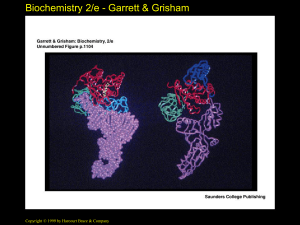
Biochemistry 2/e - Garrett & Grisham Chapter 32 The Genetic Code to accompany Biochemistry, 2/e by Reginald Garrett and Charles Grisham All rights reserved. Requests for permission to make copies of any part of the work should be mailed to: Permissions Department, Harcourt Brace & Company, 6277 Sea Harbor Drive, Orlando, Florida 32887-6777 Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Outline • • • • 32.1 Elucidating the Genetic Code 32.2 The Nature of the Genetic Code 32.3 The Second Genetic Code 32.4 Codon-Anticodon Pairing, Third-Base Degeneracy and the Wobble Hypothesis • 32.5 Codon Usage • 32.6 Nonsense Suppression Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Translating the Message How does the sequence of mRNA translate into the sequence of a protein? • What is the genetic code? • How do you translate the "four-letter code" of mRNA into the "20-letter code" of proteins? • And what are the mechanics like? There is no obvious chemical affinity between the purine and pyrimidine bases and the amino acids that make protein. • As a "way out" of this dilemma, Crick proposed "adapter molecules" - they are tRNAs! Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham The Collinearity of Gene and Protein Structures • Watson and Crick's structure for DNA, together with Sanger's demonstration that protein sequences were unique and specific, made it seem likely that DNA sequence specified protein sequence • Yanofsky provided better evidence in 1964: he showed that the relative distances between mutations in DNA were proportional to the distances between amino acid sunstitutions in E. coli tryptophan synthase Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Elucidating the Genetic Code • A triplet code is required: 43 = 64, but 42 = 16 - not enough for 20 amino acids • But is the code overlapping? • See Figure 32.2 • And is the code punctuated? Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham The Nature of the Genetic Code • A group of three bases codes for one amino acid • The code is not overlapping • The base sequence is read from a fixed starting point, with no punctuation • The code is degenerate (in most cases, each amino acid can be designated by any of several triplets Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Biochemists Break the Code • • • • • Assignment of "codons" to their respective amino acids was achieved by in vitro biochemistry Marshall Nirenberg and Heinrich Matthaei showed that poly-U produced polyphenylalanine in a cellfree solution from E. coli Poly-A gave polylysine Poly-C gave polyproline Poly-G gave polyglycine But what of others? Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Getting at the Rest of the Code • Work with nucleotide copolymers (poly (A,C), etc.), revealed some of the codes • But Marshall Nirenberg and Philip Leder cracked the entire code in 1964 • They showed that trinucleotides bound to ribosomes could direct the binding of specific aminoacyl-tRNAs (See Figure 31.6) • By using C-14 labelled amino acids with all the possible trinucleotide codes, they elucidated all 64 correspondences in the code (Table 32.3) • Read also about Khorana's experiment Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Features of the Genetic Code • All the codons have meaning: 61 specify amino acids, and the other 3 are "nonsense" or "stop" codons • The code is unambiguous - only one amino acid is indicated by each of the 61 codons • The code is degenerate - except for Trp and Met, each amino acid is coded by two or more codons • Codons representing the same or similar amino acids are similar in sequence • 2nd base pyrimidine: usually nonpolar amino acid • 2nd base purine: usually polar or charged aa Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham AA Activation for Prot. Synth. • • • • • The Aminoacyl-tRNA Synthetases Codons are recognized by aminoacyl-tRNAs Base pairing must allow the tRNA to bring its particular amino acid to the ribosome But aminoacyl-tRNAs do something else: activate the amino acid for transfer to peptide Aminoacyl-tRNA synthetases do the critical job linking the right amino acid with "cognate" tRNA Two levels of specificity - one in forming the aminoacyl adenylate and one in linking to tRNA Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Aminoacyl-tRNA Synthetases • • • • Mechanism and specificity Deacylase activity "edits" and hydrolyzes misacylated aminoacyl-tRNAs Despite common function, the synthetases are a diverse collection of enzymes Four different quaternary structures: , 2, 4 and 22 Subunits from 334 to more than 1000 residues • Two different mechanisms (See Figure 32.5) Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Recognition of tRNAs by the aminoacyl-tRNA synthetases • Anticodon region is not the only recognition site • The "inside of the L" and other regions of the tRNA molecule are also important • Read pages 1080-1082 on specificity of several aminoacyl-tRNA synthetases Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Third-Base Degeneracy and the Wobble Hypothesis • Codon-anticodon pairing is the crucial feature of the "reading of the code" • But what accounts for "degeneracy": are there 61 different anticodons, or can you get by with fewer than 61, due to lack of specificity at the third position? • Crick's Wobble Hypothesis argues for the second possibility - the first base of the anticodon (which matches the 3rd base of the codon) is referred to as the "wobble position" Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham Copyright © 1999 by Harcourt Brace & Company Biochemistry 2/e - Garrett & Grisham The Wobble Hypothesis • The first two bases of the codon make normal (canonical) H-bond pairs with the 2nd and 3rd bases of the anticodon • At the remaining position, less stringent rules apply and non-canonical pairing may occur • The rules: first base U can recognize A or G, first base G can recognize U or C, and first base I can recognize U, C or A (I comes from deamination of A) • Advantage of wobble: dissociation of tRNA from mRNA is faster and protein synthesis too Copyright © 1999 by Harcourt Brace & Company