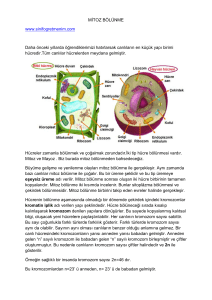
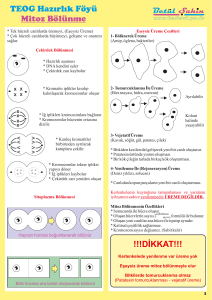
HÜCRE BÖLÜNMESİ
HÜCRE BÖLÜNMESİ
Hücreler bölünme ile sayılarını arttırırlar. Bir amip yada bakteri gibi tek hücreli hayvanlarda bölünme sonucu popülasyondaki bireylerin sayısı artar. İnsan gibi çok hücreli formlarda ise bölünme sonucu bireyi oluşturan hücrelerin sayısı artar, yani büyüme olayı meydana gelir. Bazı dokulardaki yaşlanarak yıpranan hücreler ise hücre bölünmelerinden sağlanan genç hücrelerce yenilenirler. 1. Amitoz Bölünme
2. Mitoz Bölünme
3. Meiosis (Redüksyon) Bölünme
1
HÜCRE BÖLÜNMESİ
Amitoz Bölünme
Amitoz olarak adlandırılan nukleusun doğrudan doğruya bölünmesi genellikle bazı bir hücrelilerde görülür. Çok hücrelilerde hızlı hücre çoğalması gözlenen durumlarda amitoz görülürse de daha ziyade patolojik şartlar altında ortaya çıkar. A
mitoz bölünme esnasında: sitoplazma içinde iğ iplikleri ve aster kompleksleri oluşturulmaz. Nukleus içinde kromatin materyali kromozom şeklinde belirmeden nukleus 2 parçaya ayrılır. Nukleus zarında bir parçalanma görülmez. Nukleusun ikiye ayrılmasını sitoplazmanın bölünmesi izler.
2
HÜCRE BÖLÜNMESİ
Mitoz Bölünme
Mitoz ışık mikroskobu ile gözlenebilir. Bu olay esnasında, ebeveyn hücre bölünür, ve kardeş hücrelerin her biri ebeveyn hücreninkine benzer bir kromozomal karyotip
taşır. Esas olarak, kromozomların uzunlamasına duplikasyonu gerçekleşir, ve bu kromozomlar kardeş hücrelere paylaştırılır. Bir hücrelilerden memelilere kadar tüm hayvanlarda görülen nuklear kromatin materyalinin çeşit ve miktarca eşit bir şekilde 2 yavru hücreye paylaştırılmasını sağlayan temel hücre bölünmesi tipidir. Profaz, metafaz, anafaz ve telofaz olarak adlandırılan evrelerde meydana gelir. 2 mitoz arasında hücre interfazdadır. HÜCRE BÖLÜNMESİ
Profaz: Sentrozomdaki sentriollerin duplikasyonu sonucu oluşan 2 çift sentriol
birbirlerinden uzaklaşarak hücrenin 2 ayrı kutbuna göçe başlarlar. Sentriol çiftleri
etrafında radier dağılım gösteren ve aster denilen fibriller yapılar, ve 2 aster kompleksi arasında da iğ iplikleri organize olmaya başlar. Nukleusun kromatin materyali de kromozomlar halinde belirmeye ve giderek
kısalıp kalınlaşmaya başlarlar. Her bir kromozom, birbirine paralel 2 kromatidden
ve bir sentromerden ibarettir. Profazın sonlarına doğru nuklear membran parçalanır, nukleolus belirsizleşir ve
kromozomlar sentromerleri ile iğ ipliklerine bağlanmaya başlarlar. 3
HÜCRE BÖLÜNMESİ
Metafaz
Metafaz esnasında, Nuklear zar ve nukleolus kaybolur.
Kromozomlar hücrenin ekvator düzleminde dizilirler. Kromatidler sentromer yada kinetokor bölgesinde mitotik iğ ipliklerinin
mikrotübüllerine bağlanırlar. Kromozomlar en kısa ve en kalın evrelerinde olup çift kromatid yapıları belirgindir. HÜCRE BÖLÜNMESİ
Anafaz
Anafazda, kardeş kromatidler birbirinden
ayrılıp mikrotübüllerin iğ iplikleri boyunca
hücrenin zıt kutuplarına doğru göç ederler. İmmünofluoresans iğ ipliği bölgesinde
mikrotübüler (tubulin), aktin ve myosin protein göstermektedir. Bu proteinler hücre kutuplarına kromozom
göçü sürecinde mikrotübüllere katılabilirler, fakat bu sürecin mekanizması hala tartışma
konusudur. 4
HÜCRE BÖLÜNMESİ
• Telofaz
• Bire kromatidden oluşan kardeş kromozom
gruplarının centriol kutuplarına yaklaşması ile
telofaz başlar. • Telofaz kardeş hücrelerde nukleusların tekrar ortaya
çıkmasıyla karakterize edilir. • Kromozomlar giderek belirgin şekillerini
kaybederek kromatine dönüşürler, nukleoluslar ve
nuklear membran tekrar oluşmaya başlar. • İğ iplikleri kaybolur. • Eski ekvator düzlemi boyunca iki kardeş hücreyi
sınırlayacak şekilde hücre membranlarının oluşumu
ile sitokinesis, yani sitoplazmanın bölünmesi de tamamlanıp mitoz sona erer ve yeni hücreler
interfaza geçerler
Film
5
6
HÜCRE BÖLÜNMESİ
Mitoz Bölünme
7
HÜCRE SİKLUSU
Hücre siklusu 4 evreden (profaz, metafaz, anafaz, telofaz) oluşan mitoz ve
interfaz olmak üzere 2 safhaya ayrılmaktadır. İnterfaz 3 faza ayrılır: G1 (sentez öncesi), S (DNA sentezi), ve G2 (DNA çoğalması
sonrası). DNA ile sentriollerin sentezi ve çoğalması S fazında gerçekleşir. G1 fazı esnasında, RNA ve protein sentezi gerçekleşir, ve hücre volümü normal büyüklüğüne getirilir. G2 esnasında, iğ ipliği ve aster proteinleri sentezlenir. Hücre siklusunun tamamlanması için gereken zaman hücreden hücreye ve
organizmadan organizmaya çok değişkendir. Bakterilerde genellikle 15‐20 dakika
yeterli olurken örneğin insanda 37o C de siklus ortalama 18‐24 saat gerektirir. Ancak bu süre içinde, mitoz için gereken zaman yaklaşık sadece 1 saattir. Geriye kalan süre ise interfazda, bölünme hazırlığı ile geçer
HÜCRE SİKLUSU
İğ ipliği ve aster proteinleri sentezlenir.
Mitoz (profaz,
metafaz, Anafaz, telofaz)
DNA ile
sentriollerin
sentezi ve
çoğalması
RNA ve protein sentezi
gerçekleşir, ve hücre volümü
normal büyüklüğüne getirilir
8
HÜCRE SİKLUSU
9
HÜCRE BÖLÜNMESİ
Meiosis (Redüksyon) Bölünme
Çok hücreli hayvanların gametlerinin yada eşey hücrelerinin olgunlaşması esnasında görülen ve diploid (2n) olan kromozom sayısını yarıya, yani haploide (n) indiren özel bir hücre bölünmesidir. Ard arda gelen 2 hücre bölünmesidir. 10
HÜCRE BÖLÜNMESİ
Meiosis (Redüksyon) Bölünme
Profaz I: Mayoz bölünmenin en uzun evresidir. Kromozomlar nuklear membran
içinde ince iplikçikler halinde belirir, sentrozomun sentriolleri duplike olur ve iğ iplikleri belirir. Evre ilerledikçe daha belirginleşen çift kromatidli kromozomlar homologları ile bir araya gelerek esleşir, yani bivalent yada tetrad homolog kromozom çiftleri oluştururlar. Sentromerlerinden birbirine bağlı kromozom çiftlerinin kromatidleri arasındaki chiasma denilen yakın temaslar sonucunda kromatid
parçacığı alış verişi olur. Bu olaya crossing‐over denir. Evrenin sonunda kromozomlar belirgin, nuklear membran parçalanmaya başlamış, nukleolus kaybolmuştur. Evre kendi içinde 5 ayrı alt evreye ayrılır –
Leptonema, zygonema, pachynema, diplonema ve diakinesis
11
HÜCRE BÖLÜNMESİ
Meiosis (Redüksyon) Bölünme
Metafaz I: Nuklear membran tamamen kaybolur. Bivalentler, iğ iplikleri kompleksinin ekvator düzlemine dizilirler. Her bivalentteki 2 homolog kromozomun sentromerleri ekvator düzleminin iki yanında yer alırlar. Anafaz I: Evre, homolog kromozomların birbirinden ayrılarak hücrenin zıt kutuplarına çekilme hareketleri ile başlar. Kutuplara yönelen kromozomlar tek sentromerle birbirlerine bağlı çift kromatidler halindedir ve her kromozom grubunda habloid (n) sayıda kromozom vardır. Telofaz I: Kutuplara göç eden ve belirgin şekillerini kaybetmeye başlayan kromozomlar etrafında nukleus zarlarının oluşmaya başlaması ile karakterize edilir. Yavru nukleuslar içinde nukleoluslar belirir. Çok kısa bir interfaz evresinden sonra II. meiosis bölünme başlar. 12
13
14
HÜCRE BÖLÜNMESİ
Meiosis (Redüksyon) Bölünme
Profaz II: Belirsizleşmeye başlayan kromozomlar, anafaz I’de gözlenen kromozomlarla aynı yapıdadırlar. Yani interfazda herhangi bir kromozom duplikasyonu olmamıştır. Kromozomların çift kromatidleri tek sentromere bağlı durumdadır. Nukleus zarı parçalanırken iğ iplikleri belirir. Metafaz II: Kromozomlar iğ ipliklerinin ekvator düzlemine dizilirler. Anafaz II: Her kromozomun kardeş kromatidleri iğ iplikleri boyunca zıt kutuplara doğru çekilmeye başlarlar. Bu evredeki kardeş kromozomlar birer kromatidden ibarettir. Telofaz II: Kutuplara çekilip belirsizleşmeye başlayan kromozomlar etrafında nukleus zarları oluşur, nukleoluslar belirir ve sitokinesis ile evre son bulur. Böylece, meiosis başlangıcında diploid (2n) kromozom sayılı tek ana hücreden meiosis sonunda habloid (n) kromozom sayılı 4 yavru hücre oluşmuş olur 15
GAMETOGENESIS
Dişi yada erkek bireyde gametlerin oluşması olayına gametogenesis denir. Olay, iki ayrı eşeyde birbirinden biraz farklı cereyan eder. Erkek gonadda görülene spermatogenesis, dişi gonadda görülene ise oogenesis
denir. Spermatogenesisde ilk evre, 2n kromozom sayılı primer üreme hücrelerinin mitoz bölünmelerle sayıca artmalarıdır. Meydana gelen çok sayıda diploid
kromozom sayılı hücreler spermatogonium’lardır. II. evre, her bir spermatogoniumun meiosis öncesi biraz büyüyerek gelişmesi ve primer spermatosit’leri oluşturmasıdır. III. Evrede, I. meiosis bölünme sonucu kromozom sayısı 2n’den n’e düşer, yani sekonder spermatosit’ler oluşur. Bunu hemen II. meiosis izler ve 4 adet habloid spermatid meydana gelir. Bunların değişimi ve kamçı kazanmaları ile, IV. evrede spermatozoon’lar oluşur 16
GAMETOGENESIS
Oogenesisde I. evre, ard arda gelen mitoz bölünmelerle diploid kromozom sayılı oogonium’ların oluşumudur. II. evrede, oogoniumlar büyürler ve organizma cinsine bağlı olarak, sitoplazmalarında az veya çok besin maddesi yada vitellüs toplayarak 2n yapılı primer oosit’leri meydana getirirler. III. evrede ilk meiosisi geçiren primer oosit, habloid yapıda bir sekonder oosit ve bir küçük kutup hücresi oluşturur. Hemen takibeden II. meiosis sonunda, sekonder oositten sitoplazmasının çoğunu içeren bir ootid ve küçük bir II. kutup hücresi meydana gelir. Bu arada ilk kutup hücresi de 2 yavru kutup hücresi vermiştir. IV. evrede ootid nuklear pozisyonunu biraz değiştirerek olgun bir yumurta hücresi yada ovum’u oluşturur. Diğer 3 kutup hücresi ise, zamanla çevre dokularca absorbe edilir. Yani her oogoniumdan sonuçta işe yarar bir ovum meydana gelir 17
18
19
HÜCRE’DE PROTEİN SENTEZİ
Hücrede protein sentezi DNA tarafından yönlendirilir. DNA molekülündeki belli programlara göre amino asitler polipeptid yada protein molekülleri haline getirilir. Her organizmanın kendine özgü DNA molekülleri hücre nukleusundaki
kromozomlarda yada kromatin materyalinde lokalize olmuştur. Protein sentez bölgeleri ise sitoplazmada bulunan ribozomlardır. Bu durumda şu sorular akla gelebilir. 1. Protein sentezi için gerekli bilgi DNA molekülünde nasıl şifrelenir, yada kodlanır? 2. Kod, DNA molekülünden ribozomlara nasıl aktarılır?
3. Ribosomlarda kodlara uygun protein sentezi nasıl gerçekleşir?
Sırasıyla bu sorulara cevaplamaya çalışalım.
HÜCRE’DE PROTEİN SENTEZİ
1. Şifre, DNA molekülündeki azotlu bazların belli bir düzende sıralanmaları ile oluşur. Protein sentezinde kullanılacak amino asitlerin seçimi, birbirini izleyen üçlü baz grupları halinde DNA tarafından belirlenir. Bu üçlü baz gruplarına triplet veya codon denir. DNA moleküllerinde birbirinden farklı 64 tipi olabilir (43 = 64). Bunlardan üç tanesi (UAA, UAG, UGA) anlamsız codon’lardır. Bunlara stop codon adı da verilir. Stop codonlar polipeptid zincirinin bitiş yerini belirler. Her farklı amino asit tipi genellikle birden fazla codon ile, en çok 6 farklı codon
ile şifrelenebilir. Örneğin, valin’i şifreleyebilecek codonlar: CAA, CAG, CAT ve CAC’dir. Sonuç olarak bir polipeptid molekülünde amino asit sıralanması DNA molekülü tarafından, yani genetik olarak tespit edilir
20
HÜCRE’DE PROTEİN SENTEZİ
2. DNA kodu, yani genetik enformasyon, DNA tarafından mRNA moleküllerine aktarılır. Bu olaya transcription adı verilir. Burada amaç, DNA molekülünün taşıdığı enformasyonun bir kopyasını çıkarmak ve hücrenin protein sentez bölgeleri olan ribozomlara ulaştırmaktır. Bu amaçla DNA çift zinciri açılır. Zincirlerden biri enformasyon naklinde kullanılacak mRNA molekülünü sentezler. Bu transcription olayı esnasında DNA zincirindeki Adenin, mRNA
zincirinde Uracyl ile eşleşir.
Bu mekanizmaya göre; mRNA molekülünde örneğin Valin amino asidini şifreleyecek kodonlar sırasıyla GUU, GUC, GUA ve GUG olabilecektir. Bu şekilde oluşturulan mRNA, DNA zincirinden ayrılıp sitoplazmaya geçerek bir ribozomun 40‐S parçasına yerleşir ve bu bölgeyi ribozomun 60‐S kısmına bağlar (S: Swedberg Ünitesi; Ultrasantrifüjde moleküllerin çökelme sabitesi). Ribozomda protein sentezi sona erdikten sonra mRNA çözülür. Yüksek organizmalarda yaşam süresi kısa olan mRNA tipleri yanında 1‐2 gün kalabilen mRNA’lar da rapor edilmiştir. HÜCRE’DE PROTEİN SENTEZİ
3. Tek bir mRNA molekülü protein sentezi esnasında birçok ribozomu birbirine bağlar. Bu şekilde oluşturulan yapılara poliribozom yada polysom adı verilir. Amino asitlerin bu tip ribozomlarda genetik enformasyona göre birbirlerine bağlanmalarından önce, ATP enzimlerinin aracılığı ile hücredeki kimyasal enerji devreye sokulur ve bu enerji amino asitleri aktive eder. Aktif hale geçen amino asitler sitoplazmada bulunan özel tRNA molekülleri tarafından ribozomlara taşınırlar. Amino asitleri tRNA moleküllerine bağlayan enzimlere aminoasit tRNA sentetaz adı verilir. Amino asitlerin çoğunluğu için birden fazla tRNA
mevcuttur. Tipik bir tRNA molekülünde 3 bölge ayırdedilir: 1. Amino asidin tanınma bölgesi, 2. Amino asidin bağlanma bölgesi, 3. Mesajın tanınma bölgesi ki bu bölgede anticodon grubu yer alır. tRNA molekülü ortamda tanımlamasına uygun bir aktif amino asit bulduğunda, bu amino asit tRNA molekülündeki amino asit bağlanma bölgesine aktarılır. Bu bağlantı bölgesi daima sitozin‐
sitozin‐adenin azotlu baz grubundan oluşur ve amino asit daima adenine
bağlanır 21
HÜCRE’DE PROTEİN SENTEZİ
tRNA, bağladığı amino asidi ribozomlara götürür ve antikodon tripleti ribozom üzerindeki mRNA’nın uygun bir kodon tripleti ile eşleşecek şekilde ribozoma bağlanır. 60‐S’lik ribozom ünitesi üzerinde tRNA’nın bağlanması için iki bölge vardır. Bunlardan birisi A‐Bölgesi, yani Aminoasit tRNA bağlanma bölgesi, diğeri P‐Bölgesi, yani peptidil tRNA bağlanma bölgesidir. Yeni gelen bir amino asit A‐
Bölgesine bırakılır ve P‐Bölgesine gelen bir diğer amino asit ile peptidil enzim aracılığı ile bağlanır. Bu şekilde giderek uzayan bir amino asit zinciri oluşur. Bu işlem, mRNA üzerinde bir anlamsız yada Stop Codon gelinceye kadar devam eder. Bu mekanizmaya göre ribosomlarda amino asitlerin proteinlere dönüştürülmesinden sonra ribosomlar yine alt‐ünitelerine ayrılırlar
HÜCRE’DE PROTEİN SENTEZİ
22
HÜCRE’DE PROTEİN SENTEZİ
HÜCRE’DE PROTEİN SENTEZİ
23
HÜCRE’DE PROTEİN SENTEZİ
HÜCRE’DE PROTEİN SENTEZİ
24
HAYVANSAL
HAYVANSAL DOKUDOKULAR
TİPLERİ
Epitel (Epithelial) Doku
Ektoderm
Mezoderm
Endoderm
Bağ (Connective) Doku
Mezenşim
Mezoderm
Kas (Muscle) Doku
Mezoderm
Sinir (Nervous) Doku
Ektoderm
25