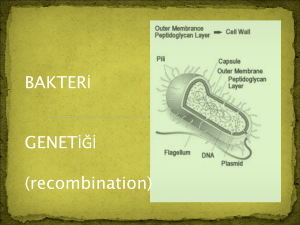
PLASMID
Mustafa EMREM
162161022
BAKTERİ GENETİĞİ
Giriş
Bazı prokaryotik (Bakteri) ve ökaryotik organizmalarda (maya, mantar, bitki),
genomik DNA’nın yanısıra, çok daha küçük, DNA karakterinde, çift iplikli
genetik elementler bulunmaktadır.
Bunlar arasında plasmidler de yer almaktadır. Ancak, bu genel tanım içine,
bakteriyofajlar, kromozomla birleşik olan profajlar, transpozonlar, İSelementleri ökaryotiklerde mitokondriumlar girmektedir
Plasmidlerin Genel Karakterleri
Plasmidler çift iplikli otonom DNA molekülleridir.
Şimdiye dek prokaryotik ve ökaryotiklerde RNA (ribonukleik asit) özelliği
taşıyan plasmidlerin varlığı bildirilmemiştir.
MA. 1-600 mega dalton, Md arasında değişmektedir.
E. coli de DNA'nın %0.04-%8'i kadardır. Büyük plasmidlere daha çok,
Gram negatif mikroorganizmalarda (Pseudomonas, Agrobacterium, vs)
rastlanılmaktadır.
Streptomyces rochei 'de iki küçük ve lineer morfolojide B. burgdorferi 'de 4-9
adet lineer ve sirküler plasmid bulunur.
Ekstrakromozomal materyaller, hücre sitoplazmasında serbest olarak bulunabilecekleri (plasmid)
gibi bakterinin kromozomu ile de birleşebilirler. Böyle plasmidlere episom adı verilmektedir.
Kointegrasyon, plasmidle genomik DNA arasında oluşabileceği gibi, plasmidle plasmid arasında
da meydana gelebilir.
Bazı plasmidler de Col E1 gibi, negatif süpersarmal (süperheliks) bir yapı özelliğine sahiptir
ve E. coli 'de 19-20 süpersarmal bulunmaktadır.
Değişik morfolojilere sahip plasmidlerin
agarose
gel
elektroforezisdeki
lokalizasyonları
ve
sentrifugasyondaki
sedimentasyon
katsayıları (S20 W) arasında farklar
vardır. Bunlar, sukroz densite gradient
(%5-20) sentrifugasyonda, santrifüj
tüpünde sukrozun değişik yoğunluğuna
sahip yerlerinde toplanırlar.
Elektroforezisde ise,
süpersarmal form karşı uçta,
lineer form ortada,
sirküler formlar ise başlangıç yerine
yakın kısımlarda yerleşirler.
Plasmidlerin kendileri ve kodladıkları spesifik informasyonların konak bakteri için hayati
önemi yoktur. Bunlar olmadan da bakteriler normal yaşamlarını sürdürürler.
Çünkü, plasmidler bakterilere sonradan transfer olmuşlardır ve özel koşullar altında da bakteriden
ayrılabilirler.
Ancak, bunlar bakterinin genomunda bulunmayan (veya bazen bulunabilen) özel karakterleri
kazandırırlar ve avantajlar sağlarlar: Örneğin,
Antibiyotikler
Metaller
İlaçlara dirençlilik
Toksin formasyonları
Pilus oluşumu
Virulens faktörleri
Fermentasyon özellikleri
Nitrojen fikzasyonu bunlardan sadece birkaç tanesidir.
Farklı özellikler kazandıran plasmidler bakterilerden ayrılırsa
veya çıkarılırsa (plasmid eliminasyonu, plasmid eksklusyonu,
curing), bakteriler kazandıkları bu avantajları kaybederler ve bu
yönlerden negatif hale gelirler.
Örn, C. tetani 'de toksin formasyonu bir plasmid tarafından
kodlanır. Bu plasmidin çıkması nonpatojenik hale getirir.
İn vitro koşullarda ve amaca uygun olarak kendilerine istenilen
özel markerler eklenerek hazırlanan suni plasmidler (pBR322,
pUC8/9 vb.) rekombinant DNA teknolojisinde, genlerin
klonlanmasında aracı molekül (vektör) olarak daha fazla
kullanılmaktadırlar.
Genellikle küçük veya orta boy suni plasmidlerden daha fazla yarar
sağlanmaktadır, kromozomla birleşmediğinden veya sitoplasmada yeterince
bulunduklarından, hücrelerden çıkarılmaları oldukça kolaydır.
Büyük plasmidler, hem kendilerini ve hem de taşıdıkları informasyonları ve
konakçı kromozomundan bir geni, kendine yakın türlere transfer etme yeteneğine
sahiptirler.
Buna karşın, küçük plasmidler ise, ya aynı bakterideki büyük plasmidlerin yardımı
ile veya konakçının genomu ile birleşerek bakteri bölünmesi sırasında kardeş
hücrelere transfer edilirler.
Bunlardan ayrı olarak, genel transdüksiyonlada
küçük plasmidler transfer olabilirler.
Kimyasal ve fiziksel yöntemler kullanarak plasmidlerin bakteriden çıkması
kolaylaştırılır ve serbest hale getirilebilirler.
Konjugatif plasmidlerde bulunan seks pilusu formasyonunu kodlayan
genler, nonkonjugatiflerde bulunmamaktadır.
Bakteri DNA'sının replikasyonuna mani olan mutasyonlar aynı
zamanda plasmid replikasyonunu da bloke ederler.
Küçük plasmidler bakteri kromozomu ile senkronize replike
olmadıkları için böyle mutasyonlardan etkilenmezler. Bunlar ancak,
DNA pol I'de oluşan mutasyonlara duyarlıdırlar.
Plasmidlerin Yapısı
Plasmidler, genellikle, konakçı bakteri içinde kendi bağımsız durumlarını
koruyabilecek ve devam ettirebilecek gerekli genetik informasyonlara sahip
otonom ünitelerdir.
Plasmidlerde bulunması gerekli olan en önemli sekansların başında,
replikasyonda fonksiyonu olan replikasyon orijini (ORI)'ni kodlayan
nukleotidlerdir.
ORI'ya sahip olan küçük veya büyük DNA sekansları bağımsız replikasyon
özelliği gösterebilirler (replikon). Replikasyon orijini plasmidlerde genellikle tek
olmasına karşın, bazılarında R6K plasmidinde 2-3 ORI bulunabilmektedir.
F plasmidleri : ORI + pilus formasyon genler
R
plasmidleri
:
ORI
genleri+antibiyotiklere dirençlilik
Virulens plasmidleri : ORI + toksin genleri (B. anthracis 'te p0X1
ve Cl. teteni 'de nörotoksin)
Bakteriosin plasmidleri : ORI + Co1E1, Col I, Col V, Pesticin,
Cloacin, Megacin vs. genler
Metabolik plasmidler : ORI + bazı kimyasal maddeleri (SAL,
ASL, TOL, NAH, vs) ayrıştıran genler
+
genleri
pilus
formasyon
Plasmidler, spesifik restriksiyon endonuklease enzimlerinin
kesim yerlerine ve restriksiyon haritalarına sahiptirler.
Enzimler, türlerine göre, plasmidlerde farklı sayıda kesmeler
yaparak değişik boylarda DNA segmentlerine ayırırlar.
Bazı enzimlerin (Bst II, Hpa I ve Xba I) plasmidde (pBR322) hiç
bir kesim yeri bulunmamaktadır.
Buna karşın, Hpa II'nin bu plasmidde 26 yerden kesme yeteneği
vardır.
Sadece bir kesim yerinin bulunması ve spesifik markerlerden
birinin üzerinde olması, genin inaktivasyonu için istenilen bir
durumdur (EcoR I, Pst I, gibi).
Aşağıda, bazı suni plasmidlerde amaca yönelik olarak ilave edilen bazı spesifik
markerler gösterilmektedir.
pMB 9 (5300 bp): ORI + tetr
pBR322 (4363 bp): ORI + ampr+ tetr
pNEO (5510 bp): ORI + ampr+ neor
pBR 325 (6000 bp) : ORI + ampr + Camr+ tetr
pUC 8/9 (2655 bp): ORI + ampr+lac
04.
Plasmidlerin Sınıflandırılması
Bakteri plasmidleri bazı klasifikasyonlara tabi tutulmuştur. Ancak, yine de tam yerleşmiş ve
bütün kriterlere uyabilecek veya cevap verebilecek bir sınıflama yapılmış değildir.
1) Büyüklüklerine göre sınıflama: Bazı araştırmacılar plasmidleri büyüklüklerine veya molekül
ağırlıklarına göre 3 gruba ayırmaktadırlar,
a) Küçük (mini sirküler) plasmidler: Bunlar 1-10 x106 dalton ağırlığındadırlar
b) Orta plasmidler: Bunlar, 10-50x106 molekül ağırlığındadır.
c) Büyük plasmidler: Bu ölçülerin üstünde molekül ağırlığına sahip olanlar.
2) İnkompatibilite durumlarına göre sınıflama: Birden fazla plasmid aynı bakteride uyum içinde
bulunabiliyorsa bunlar kompatible uyum içinde olamıyor ve birbirlerini itiyorlarsa inkompatible kabul
edilmektedirler. Birbirlerinin üremelerine veya diğerinin replikasyonuna da mani olmaktadır.
3) Konjugatif özelliklerine göre sınıflama: Küçük (1-10 kbp uzunlukta) plasmidlerin transferi, aynı
bakteri içinde bulunan diğer konjugatif plasmidlerce sağlanır veya bakteri kromozomu ile birleşerek
veya genel transdüksiyonla gerçekleştirilir. Bu tür plasmidler, yapılarında seks pilusu formasyonu için
de genetik kodlar taşımazlar.
4) Taşıdıkları spesifik sekanslara göre sınıflama: DNA'larında bazı özel markerler kodlarına
sahiplerdir. Konjugatif plasmidler bu özelliklerini transfer edebilme yeteneğindedirler. Bu markerlere
göre plasmidler 5 gruba ayrılırlar: F faktörü (seks pilusu), Rezistenslik plasmidleri, Virulens
plasmidleri, Bakteriyosin plasmidleri, Metabolik plasmidler
05. Plasmidlerin Bakteride Saptanması
Bakteriyel populasyonlarda plasmidlerin varlığını belirlemede fiziksel, morfolojik ve fonksiyonel
testlerden yararlanılmaktadır.
Bakterilerin, bazı özel karakterleri çabuk kazanması ve bunları nesillere transfer etmesi veya bunları
çabuk kaybetmesi, bu özelliklerin plasmidlerce kodlandığının bir işareti olarak kabul edilmektedir.
Elektroforezisde Plasmid DNA'sı karşı uçta yer alırken, büyük olan kromozomal DNA başlangıç yerine
yakın kalır.
Elekron mikroskopuyla plasmid DNA'sı kolayca görüntülenebilir. Plasmid DNA'sı kromozomal DNA'dan,
büyüklük farkı nedeniyle kolayca ayırt edilebilir.
Nukleusla birleşenleri saptamak için spesifik işaretli problardan yararlanılabilir.
Ethidium bromide caesium chloride densite gradient sentrifugasyona tabi tutularak sirküler plasmid
DNA'sı dibe yakın yerde, kromozomal DNA ise bunun üstünde bant oluşturur.
06.
Plasmid DNA'sının İzolasyonu
Bakterinin içinde plasmidin varlığı saptandıktan sonra, kontamine olmadan plasmidleri çıkarmak ve bazı özelliklerini
belirlemek mümkündür. Bu amaç için, genellikle, sitoplasmada bulunan plasmidler tercih edilirler.
1)Rapid boiling method for small plasmids in E. coli (Holmes & Quigley,1981; modified by Riggs & McLachlan, 1986)
2)Hot alkaline method for all plasmid sizes and bacteria (Kado & Liu,1981), modified
3) Lysozyme method for various Gram-negative bacteria (Davis et al.,1980)
4) Lysis of cells from single colonies on agarose gel (Eckhardt, 1978; Priefer, 1984), modified
5)Plasmid isolation from Gram-positive bacteria, especially lactobacilli, with mutanolysin or lysozyme (Klaenhammer,
1984)
6) Isolation procedure for all plasmid sizes from all bacteria (Crosa et al., 1994)
07. Plasmid Eliminasyonu (Curing, Plasmid Eksklusyonu)
Bakteriye ait özelliklerin büyük bir kısmı kromozomal orijinli diğerleri ise plasmidler tarafından
kodlanmaktadırlar. Bu iki durumu birbirinden ayırmak için plasmid eliminasyon testi yapılır.
Plasmid eliminasyon testinde, genellikle, kimyasal maddeler (acridin orange, acriflavine, ethidium bromide,
mitomycin C,optimalin üstündeki ısı derecelerinde üretme, UV-ışınları, iyonizan ışınlar, vs) fazla kullanılır.
Plasmidlerin bakteriden ayrıldığını anlamak için ise İndikatör mikroorganizma, plasmidlerin taşıdığı yeni
karakterleri alarak pozitif hale gelir. Ortama CaCl2 katılarak permeabilitesi arttırılır ve böylece transfeksiyon
kolaylaştırılır. Bu karakterler, dirençlilik veya metabolik plasmidler, vs olabilir.
Diğer teknik ise, selektif ortamların kullanılmasıdır. Özellikle, antibiyotiklere duyarlılığın oluştuğunu veya
fermentasyon durumunu ortaya koyabilen özel besi yerleri bu amaç için kullanılabilir.
08. Plasmid Replikasyonu
Plasmidler, hücre içinde, aynen bir bakteri kromozomu gibi replikasyon modeline sahiptirler. Üzerinde en fazla
çalışılan ColE 1 plasmidin replikasyonudur ve unidireksiyonaldır
R-plasmidlerinden olan R1, R100 ve R6-5'de unidireksiyonaldır
E. coli 'ye ait bazı plasmidlerin replikasyonu iki yönlü olabilmektedir (bidireksiyonal). Bu model replikasyon,
bakteri kromozomunun sentezine aynen uymaktadır. Replikasyon çatalı iki yönde ilerleyerek belli bir noktada
birbirleriyle birleşirler.
Plasmidlerde replikasyon orijini genellikle, tek olmasına karşın bazı F-plasmidinde iki ve R6K'de ise 3 tane
RO'nun bulunduğu açıklanmıştır.
09. İnkompatibilite
Bir bakteride bulunan replikasyon mekanizması bir plasmid tarafından kullanılıyor ve bloke ediliyorsa,
aynı mekanizmadan yararlanan ikinci plasmid bu olanağa sahip olamadığı için replike olamaz
(kompetatif inhibisyon).
Bu durum aynı zamanda, aynı plasmid ile ikinci infeksiyona da mani olur (immunite).
E. coli 'ye ait olan F plasmidi sitoplasmada serbest olarak veya bakteri kromozomu ile birleşik olarak
(Hfr hücre) bulunur. İki tarz birlikte E. coli 'de bulunamaz. Böylece biri elimine edilir. Genellikle, aynı
tür plasmidler birbirleri için inkompatibil bir karakter taşırlar.
10.Nonkonjugatif Plasmidlerin Transferi
Konjugatif plasmidler kendilerini transfer edebilir ancak nonkonjugatif olanların böyle bir yetenekleri
bulunmamaktadır.
Konjugatifler, kendilerini transfer ettikleri oranda, nonkonjugatifleri de mobilize ederek transferlerini
sağlayabilirler.
Mobilizasyon bazı durumlarda, konjugatif plasmide bağlanarak bununla birlikte (kotransfer) ve ne de
bakterinin recA gen ürünü proteinine bağımlıdır. Kendilerinin mobilizasyonunu sağlayan konjugatif
plasmidler bu amaç için en az bir proteinin kodlarına sahiptirler.
11. Plasmid Profil Analizi (Plasmid Fingerprint Testi)
Bu test, bakterilerde plasmidlerin varlığını ortaya koyduğu gibi plasmid türlerini belirlemede de yardımcı
olmaktadır.
Bakterilerde aynı molekül ağırlığında olanlarla farklı olanların fingerprintleri, agarose jel üzerinde farklı
yerlerde lokalizasyon gösterdiklerinden, bu durumları teşhiste büyük kolaylıklar sağlamaktadır.
12.1.
F plasmid (F Faktörü, Seks Faktörü)
F faktörü (fertilite faktörü) en iyi E. coli K 12 da incelenmiştir.
Bu plasmid, yapısında replikasyon orijininden ayrı olarak kendisinin transferini sağlayabilecek ve pilus
formasyonunu kodlayan bilgilere sahiptir (Tra operonda 21 gen).
Bu plasmid hangi hücreye aktarılırsa, o bakteride hücre membranında pilus sentezi meydana gelir.
Bu pilus (seks pilusu), bu faktörü taşımayan alıcı hücrenin (F- hücre), verici hücre ile (F+ hücre) bağlantı
kurmasına ve bir konjugasyon köprüsü oluşturmasına yardımcı olur. F faktörü konjugatif (transmissible) bir
plasmiddir.
F faktörü bakterinin sitoplasmasında bulunabileceği gibi, konakçının genomuna da integre olabilir ve Hfr
(high frequency recombination) hücre meydana gelir.
F plasmidi ile başlıca 3 tür konjugasyon oluşabilmektedir.
1) F+ hücre x F- hücre konjugasyonu: F+ hücre ile
buna sahip olmayan F- hücre arasında gerçekleşen F
faktörü aktarılmasında seks pilusu önemli fonksiyona
sahiptir. F+ hücre x F¯ hücre birleşmelerinde genin
aktarımı tek yönlüdür
2) Hfr hücre x F-hücre konjugasyonu: Alıcı hücreye
aktarılan F faktörü veya F faktörü + kromozom segmenti,
ya alıcı hücrenin genomuna integre olur ve bu hücreyi
de Hfr haline getirir veya F plasmidi, hücre
sitoplasmasında da kalabilir.
3) F' hücre x F- hücre konjugasyonu: Hfr hücrelerinde
kromozomda yerleşik bulunan F plasmidi buradan
ayrılabilir. Bu ayrılma tam veya kısmi olabildiği gibi bazı
durumlarda da kromozomdan da bir segmentin veya
genin birlikte çıkmasına neden olabilir.
12.2. Rezistenslik Plasmidleri (R Plasmidleri)
Enterobakteriler arasında çeşitli ilaçlara karşı dirençlilik, hem in vitro ve hem in vivo olarak saptanmıştır. İlk
defa bu özellik, 1951 yılında Japonya'da seyreden bir dizanteri olgusundan izole edilen S. flexneri suşunun
4 antimikrobial maddeye (kloramfenikol, tetrasiklin, streptomisin ve sulfonamid) karşı dirençli olduğu
R plasmidleri çift iplikçikli sarmal, sirküler ve yapıda DNA karakterinde genetik elementlerdir. Genellikle
konjugatif bir özelliktedirler Bakteri kromozomu ile de integre olabilirler.
R faktörleri iki bölümden oluşurlar. Biri konjugasyonu ve aktarılmayı yöneten transfer faktörü (TF) ve
diğeri de çeşitli ilaçlara karşı dirençliliği tayin eden rezistenslik faktörü (RF) veya R determinanttır.
E. coli 'de bu iki faktör tek bir ünite halinde bulunmasına karşılık salmonella ve proteuslarda bunlar ayrı
sekanslar halindedirler. Bu faktörler birbirlerinden bağımsız olarak aktarılabilirler ve akridin boyalarından
da ayrı ayrı etkilenirler.
Eğer, RF transfer edilirse alıcı hücre ilaçlara karşı dirençli hale gelir. Ancak, RF'nin, TF olmadan tek başına
aktarılması olanaksızdır.
12.3. Bakteriyosin Plasmidleri
Bakteriyosinler, ancak kendini sentezleyen türden veya konakçısına çok yakın olan
mikroorganizmalara etkilidirler.
Örn., E. coli tarafından sentezlenen colicin bunu sentezleyemeyen E. coli 'ler ile S.
sonnei için letaldir.
Şimdiye kadar 20'den fazla ve ayrı özellikte kolisin izole edilmiş olup bunlar A'dan V'ye
kadar, büyük harflerle bir sıralamaya tabi tutulmuştur.
Bunların her biri, E. coli 'lerin bazılarına etkili olmakta ve böylece bir konakçı spektrumu
ve spesifitesi meydana gelmektedir.
Bakteriyosinler, Gram negatif mikroorganizmaların yanı sıra bazı Gram pozitiflerce de
(stafilokok, streptokok, listeria, basiller vs) sentezlenebildikleri görülmektedir
12.4. Virulens Plasmidleri
Patojenik mikroorganizmalar, kendilerinde bulunan bazı virulens faktörleri ile konakçıyı hastalandırırlar ve
hatta ölümlerine neden olabilirler.
Bakteri, ne kadar virulent olursa olsun, eğer konakçı buna direnç gösteriyorsa, mikroorganizma üreyemez,
infeksiyon veya hastalık oluşturamaz.
Bakterilerin sahip oldukları virulens faktörlerinin bir kısmı da kromozomlarında kodlanmasına karşın bazıları
da plasmid orijinlidir.
C. tetani (nörotoksin), B. anthracis (pOX1 letal toksin), B. thuringiensis(kristal toksin, insekt larvaları için
toksik) A. tumefaciens (dikotiledon bitkilerde tümör formasyonu), L. lactis (nisin), S. aureus (eksfoliatif
toksin)
12.5. Metabolik Plasmidler
Metabolik aktivite bir çok bakteride kromozomlarda bulunan özel genler tarafından yönetilirler. Ancak, bazı bakterilerde
de metabolizma ile ilgili reaksiyonlar hem kromozomal ve/veya hem de plasmid orijinli olabilmektedir.
1) Degradatif plasmidler: P. putida ve diğer pseudomonaslarda bir çok kimyasal maddeyi ayrıştırabilecek plasmid
kaynaklı enzimlere sahiptirler. Böyle enzimlerin sayısı 10'dan fazlayı bulmaktadır. Örn., SAL, NAH, ASL ve TOL
plasmidleri, sıra ile salycylate, naphtalene, alkyl benzen sulphonate ve tolueni ayrıştırarak catechola çevrilir ve sonra da
plasmidlerce kodlanan enzimler tarafından acetaldehyde ve piruvatlara dönüştürülür.
2) Diğer metabolik plasmidler: Salmonellalar, genellikle, Lac-'dirler. Bazen Lac+ fenotipe dönüşebilirler (S. typhi).
Bunları, Lac+ plasmidler gerçekleştirirler. Lac+ plasmidlere, salmonellalar dışında, serratia, S. lactis ve proteuslarda da
tesadüf edilmiştir. Plasmidler, aynı zamanda, salmonellalarda, sukroz fermentasyonuna da neden olabilirler.
12.6. Büyük Plasmidler
Genel bir kural olmamakla beraber, molekül ağırlığı 50 Md'den fazla olanlar "büyük
plasmidler" olarak dikkate alınmaktadırlar.
Gen aktarımlarında önemli fonksiyonları olan plasmidlere, kirli ve organik materyaller
açısından zengin sularda yaşayan bakterilerde rastlanılmaktadır.
13. Antimikrobiyallere ve Metallere Dirençlilikte Plasmidlerin Rolü
1) Antibiyotiklerin etkilediği hedef bölgelerin modifikasyonu,
2) Antibiyotikler modifiye edilerek etkinliğinin inaktivasyonu,
3) Antibiyotiklerin hücreye girişinin önlenmesi
4) Enzim substitusyonu
13.1 Hedef Bölgenin Modifikasyonu Stafilokok ve streptokok türlerinde bulunan plasmidlerin kodladığı enzimler,
bakteriyel ribosomlardaki 23S rRNA da bulunan iki adenin molekülünü metile ederek, lincomycin ve
erythromycinin bunlara bağlanmasını önlerler.
13.2 Antibiyotiklerin Modifikasyonu Aminoglikozidlere (streptomycin, spectinomycin, kanamycin, gentamycin,
amikacin) karşı dirençliliklere karşı antibiyotikleri modifiye eden özel enzimlerin rolü fazladır. Modifiye olan
antibiyotikler ribosomlara bağlanamazlar.
13.3. Antibiyotiklerin Girişinin Önlenmesi Enterobakterilerde rastlanan Tn10 (Transposon 10) tetrasiklinlere
dirençlilikte önemli fonksiyona sahiptir. Plasmidlerin sağladığı rezistenslikte tetrasiklinlerin hücreye girişi önlenir
ve dışarı atılır.
13.4. Enzim Substitusyonu Sulfonamidler, dihidroptoreate sentezinde fonksiyonu olan enzimin kompetatif
inhibitörleridirler. Bu enzim (dihidroptoreate sentase), dihidrofolatın sentezi için gereklidir.
13.5. Metallere Dirençlilik Enterobakterilerde ve stafilokoklarda Arsenik, gümüş, bakır, cıva ve tellurium
iyonlarına, stafilokoklardaki plasmidler, arsenik, bizmut, kadmium, bakır, kurşun, civa ve çinko bileşiklerine,
Pseudomonaslardaki plasmidler de krom, civa ve tellurium iyonlarına karşı dirençlilik mekanizması oluştururlar.
PBR322 PLAZMİTİ
pBR322 -1977’de Herbert Boyer,
California Ünv. lab
p, plazmit
BR, Post-doc "Bolivar" ve "Rodriguez”
PBR322 PLAZMİTİ
Ampisilin ve tetrasiklin
direnç genleri
Birçok restriksiyon enzim
bölgesi
PUC PLAZMİTİ
pBR322’nin yarısı büyüklüğünde
(daha büyük DNA’ların klonlar)
pBR322’den geliştirildi
•
Messing et al.
•
p, plazmit
•
UC, University of California
•
pUC18, pUC19’a benzer, MCS ters