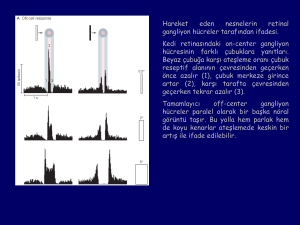
Hareket, Derinlik ve
Form Algısı
* Hareket ve derinlik algısı dorsal yolakla
posterior pariyetal kortekse
* Form ve renk algısı ventral yolakla inferior
temporal kortekse
Sitokrom oksidaz boyaları V1’deki
blobları V2’deki ince ve kalın
stripeları ortaya çıkarır.
V1’deki bloblar ve interbloblar
sırasıyla V2’deki ince stripelara ve
interstripelara bağlanır V4’e
projeksiyon temporo-oksipital
korteks inferior temporal korteks
4B tabakası V2’deki kalın stripelara
projekte olarak veya doğrudan
Middle temporal alana projekte olur.
Pariyetal yolak primer olarak M yolağından input alır; temporal yolak ise hem M hem de P
yolağından input alır.
V1’de hem magno hem de parvoselüler yolakların “bloblara” girdisi var. V2’de “stripe”
kompartmanları arasında karşılıklı bağlantılar var.
LGN’un magnoselüler tabakalarının blokajı MT ve V4 hücrelerinin aktivitesi azalır.
parvoselüler tabakalarının blokajı MT’de zayıf bir etki oluşturur. V4 aktivitesi azalır.
Retina, LGN ve hatta striat ve ekstrastriat korteksin birçok
bölgesindeki nöron reseptif alanı boyunca hareket eden bir
ışık noktasına çok iyi yanıt verir.
V1 alanındaki hücreler belirli bir doğrultudaki harekete yanıt
verirler. Bu doğrultusal seçicilik 4B tabakasında belirgindir.
M yolağındaki hücreler 4B’deki hücrelere girdi sağlar, ama
kendileri doğrultusal seçicilik göstermez.
Nöronun hareketin doğrultusuna seçiciliği
presinaptik nöronların yanıt latanslarına
bağlıdır. “a” ve “b” nöronlarının uyaranın
başlangıcına göre yanıt latansları “d” ve “e”
nöronlarına göre uzundur. Uyaran soldan
sağa hareket edince ilk olarak “a” ve “b”
nöronları uyarılır ama yanıtları geciktiği için
“d” ve “e” nöron girdileri ile aynı anda ulaşır
ve hedef hücrede toplanarak ateşlemesine
neden olurlar.
Maymunda middle temporal alandaki hemen tüm hücreler
doğrultuya seçicidir. MT alan da kontralateral görme
alanının retinotopik haritasını içerir, ama hücrelerin reseptif
alanı striat korteksteki hücrelerden 10 kat daha geniştir.
Benzer doğrultusal seçiciliği olan hücreler vertikal
kolonlarda organize olmuştur. Görme alanının her bir
bölümü farklı doğrultudaki harekete yanıt veren hücrelerden
oluşan bir grup kolon ile reprezante edilir (V1’deki
kolumnar organizasyona benzer).
İnsanda hareket ile ilgili bölge pariyetal,
temporal ve oksipital korteksin kesişim
bölgesindedir.
Denekler hareket halindeki noktalardan
oluşan bir paterne bakarken elde edilen
PET görüntüleri.
Üç grating (ızgara) üç farklı
doğrultuda
hareket
ediyor.
Küçük dairesel bir delikten
bakıldığında her üçü de aynı
doğrultuda (aşağıya ve sağa)
hareket ediyor gibi görünür. V1
ve MT alandaki çoğu nöronun
reseptif alanı küçük olduğundan
hareketin gerçek doğrultusunu
tespit etmede yetersiz kalırlar.
Kendi oryantasyon eksenlerine dik doğrultudaki harekete yanıt veren iki veya
daha fazla hücre farklı doğrultularda hareket eden farklı kenarlar ile uyarılırlar.
Yüksek-seviyeli bir hücre düşük-seviyeli hücrelerden gelen sinyalleri birleştirerek
nesnenin tümünün hareketini kodlayabilir.
Bu hipoteze göre iki evreli bir süreç vardır:
V1 ve MT alandaki bileşenin doğrultusuna seçici
nöronlardan MT alandaki paternin doğrultusuna duyarlı
nöronlara (%20 kadarı) gelen inputlar ile bir nesnenin
global hareketi hesaplanır.
MT nöronlarının mikrostimulasyonu ile hareketin algılanan doğrultusu
değişir:
Maymuna %25 korelasyona sahip hareket
eden noktalar uygulandığında ve hareketin
doğrultusunu belirtmesi istendiğinde her
bir doğrultu için eşit olasılıkta seçim yapar.
Belirli bir doğrultudaki (225°) harekete en
iyi yanıt veren hücrelerin oluşturduğu
kolon elektriksel olarak uyarıldığında (1 sn
süreli ve görsel uyaranın başlama ve bitişi
ile senkron) maymun daha çok hareketin o
doğrultuya doğru olduğu yönünde karar
verir .
Derinlik algısı
1. Monoküler ipuçlarına
2. Binoküler eşleşmemeye bağlıdır.
Monoküler ipuçları uzak alan derinlik algısını yaratır
Yüz feet (30 m) üstü uzaklıklarda
her iki gözün gördüğü retinal
görüntüler aynıdır. Buna rağmen
monoküler derinlik ipuçları ile tek
gözle derinliği algılayabiliriz.
Oklüzyon; bir nesne diğerini kısmen
örtüyorsa öndekinin daha yakında
olduğunu düşünürüz. Doğrusal
perspektif; paralel çizgiler uzaklıkla
birbirine yaklaşır. Ölçü perspektifi;
iki benzer nesne farklı ölçülerde
görünüyorsa küçük olanın daha
uzak olduğu düşünülür.
Ölçüye aşinalık; deneyimimiz ile bir nesnenin ölçüsünü biliyorsak o nesnenin
uzaklığını kestirebiliriz. Hareket; başımızı veya vücudumuzu bir taraftan diğer
tarafa hareket ettirdiğimizde baktığımız nesneden daha yakın olan nesneler daha
hızlı ve hareketimize ters doğrultuda, daha uzak olanlar daha yavaş ve
hareketimizle aynı doğrultuda hareket ediyor gibi görünür.
Stereoskopik ipuçları yakın alan derinlik algısını yaratır
Her bir göz dünyayı hafifçe farklı pozisyonlarda görür.
Farklı uzaklıklardaki nesneler iki retinada hafifçe
farklı görüntüler oluşturur. Bir noktaya odaklanıldığı
zaman noktanın görüntüsü her bir gözde retinanın
merkezindeki “eş” noktalara düşer. Derinlik için ipucu
fiksasyon noktasının hemen proksimal veya
distalindeki noktalar aracılığı ile sağlanır. Bu noktalar
her bir gözün retinasında hafifçe farklı kısımları
uyararak binoküler eşleşmeme (binocular disparity)
oluştururlar. Görüntünün iki gözün merkezinden
uzaklığı aracılığı ile nesnenin fiksasyon noktasına
uzaklığı hesaplanır.
Nesnenin bize daha yakın olan bölümleri iki retinada
horizontal doğrultuda birbirinden uzak bölgelere; daha
uzak bölümleri ise iki retinada birbirine daha yakın
bölgelere projekte olurlar.
İki göz dünyaya hafifçe farklı açılardan baktığı için
fiksasyon düzleminin önünde veya arkasında uzanan
nesneler iki retinada eşleşmeyen noktalara projekte
olurlar.
Eşleşmeme küçük ise görüntüler birleşir ve eşleşmeme
derinlikte küçük farklılıklar olarak algılanır.
Eşleşmeme daha büyük ise çift görme oluşur (bu
normal fenomen genellikle farkedilmez).
İki gözden bilgi ilk olarak primer görsel kortekste birleştirilir
Stereopsis için iki retinal görüntüde horizontal bir
eşleşmeme olmalı. P noktasına bakıldığında görüntüler
iki retinada üst üste gelir, binoküler eşleşmeme sıfırdır.
Q noktasındaki görüntü sol gözde aynı noktaya düşer
ama sağ gözde lateral olarak yer değiştirmiştir,
binoküler eşleşmeme vardır. V1’deki belirli nöronlar
horizontal eşleşmemeye seçicidir.
Binoküler girdi alan bazı kortikal nöronlar her iki
gözden gelen gidilerin eşleşmemesi sıfır olduğu zaman
(monoküler reseptif alanları görsel uzayın aynı
bölgesini örnekleyen binoküler hücreler); bazıları ise
her iki gözden gelen girdiler iki retinada uzaysal
olarak eşleşmediği zaman maksimal olarak aktiftir.
V1’e ek olarak V2 ve V3 ekstrastriat alanlarındaki ve
MT alandaki çoğu doğrultuya-seçici hücre belirli
uzaklıklardaki uyaranlara en iyi yanıt verir.
Ventral kortikal yolak form ve renk analizi ile ilgilidir
V1’den V2 ve V4’e oradan da inferior temporal kortekse gider.
V2’nin ince ve interstripeları V4’e projekte olur. V2’deki hücreler
uyaranın oryantasyonuna, rengine, horizontal eşleşmemesine
duyarlıdır ve V1’de başlayan kontur analizini sürdürürler.
V4’teki hücrelerin bir kısmı renk ve formun kombinasyonlarına yanıt
verirler.
Yüzlerin ve diğer kompleks formların tanınması inferior temporal
korteks süreçlerine bağlıdır. Hücrelerin tümünün reseptif alanı foveal
bölgeyi içerir; çok geniştir (bazen her iki görme yarı alanı); belirli bir
retinotopik organizasyonları yoktur.
İnferior temporal korteks hücreleri hem şekle hem de renge duyarlıdır.
Çoğu hücre çeşitli şekil ve renklere yanıt verir. Ama yanıtın gücü şekil ve
rengin değişik kombinasyonları için farklıdır. A. Farklı şekillerdeki
uyaranlara tek bir nöronun ortalama yanıtı (ateşleme oranı). B. Renkli
uyaranlara aynı nöronun yanıtı. Ateşleme oranı dairenin boyutu ile
gösterilmiş.
Bazı inferotemporal hücreler yalnız yüz veya el gibi spesifik uyaranlara
yanıt verirler. Yüz için seçici olan nöronların bir kısmı yüzün önden
diğerleri ise yandan görünüşüne en duyarlıdır. Bazı nöronlar seçici olarak
yüzlere bazıları ise spesifik yüz ifadelerine yanıt verirler.
İnferior temporal korteksteki bir nöronun yanıtı. Hücre oyuncak bir
maymunun yüzüne yanıt verir. Yanıtı oluşturan kritik özellikler gri bir
daire üzerinde iki siyah nokta ve horizontal bir çizgi. Dairenin içi ve dışı
arasındaki kontrast önemli değil. Ama noktalar ve çizgi dairenin içinden
daha koyu olmalı.
İnferior temporal korteks nesne
tanımaya katılan kortikal ağın bir
parçasıdır: Nesne tanıma, görsel
sınıflama, görsel bellek ve duygu
ile iç içedir.
Mediyal temporal lob
Prefrontal korteks
Amigdala
Superior temporal polisensory alan
Nesne tanıma “algısal tutarlılığa”
dayanır. Nesnenin boyut, pozisyon,
rotasyon gibi basit dönüşümleri
karşısında algının değişmemesi.
Farklı uzaklıklardaki bir nesne
retinada farklı ölçülerde görüntü
oluştursa da aynı olarak algılanır,
boyut tutarlılığı. İnferior temporal
nöronların seçici yanıt paterni
uyaran boyutu ile değişmez.
İnferior temporal korteksteki tek bir
hücreden kayıt
Retinal
görüntüdeki
pozisyon
değişikliklerine rağmen nesne aynı
olarak algılanır, pozisyon tutarlılığı.
İnferior temporal nöronlar görme
alanının farklı yerleşimlerindeki
aynı uyarana benzer yanıt verir.
Algısal tutarlılık, görsel sistemin tek bir nesne tarafından
oluşturulan farklı retinal görüntüleri genellemesidir. Daha
genel bir tip tutarlılık nesneleri kategorik olarak
algılamaktır. Kategorik algı davranışı kolaylaştırır.
Earl Miller ve ark. belirli bir kategorideki nesnelere aynı,
farklı kategoridekilere farklı yanıt veren bir nöron grubu
var mı?
Görüntülerde kedi ve köpek özellikleri farklı oranlarda birleştirilmiş.
İnferior temporal korteksten doğrudan yanıt alan lateral prefrontal korteksteki
görsel nöronlardan kayıt. Nöron köpek görüntülerine seçici yanıt vermiş. Farklı
retinal görüntülere rağmen her bir kategorideki imajlara yanıt benzer. Farklı
kategorideki görüntülere yanıtlar ise farklı.
Görsel sistemdeki birleştirme (“binding”) problemi
Ayrı olarak işlenen farklı tipteki görsel
bilginin tek bir görüntü oluşturmak
üzere birleştirilmesi için önerilen bir
model.
Farklı özellikler paralel olarak farklı
beyin bölgelerindeki farklı özellik
haritalarında kodlanır. Master haritası
özellik haritalarından girdi alır ama
yalnız
dikkat
edilen
nesneyi
çevresinden ayıran özellikleri tutar.
Dikkatin, özellikleri birbirine bağlayıcı
etkisinin olduğu öne sürülmüştür.
(Anne Treisman & Bela Julesz).
Bir nesne görme korteksindeki bir grup nöronu aktiflediği zaman
nöronlar birlikte ateşlemeye ve osilasyon yapmaya eğilim
gösterirler. Osilasyonların hücreler arasındaki senkronluğun bir
göstergesi olduğu ve aynı nesnenin farklı özelliklerine yanıt veren
hücrelerin aktivitelerini biraraya getirdiği düşünülmüştür.
(Charles Gray, Wolgang Singer & Reinhold Eckhorn ve ark.).
Nöronlar arasındaki senkronizasyon farklı kortikal alanlardaki
nöronlara yayılmalı.
LGN’dan inferior temporal kortekse projekte olan nöronların
ateşlemelerinin temporal paterni daha fazla bilgi taşır.
Ateşlemelerin temporal dağılımı değerlendirilerek uyaranın farklı
özelliklerinin aynı hücrenin yanıt paternindeki farklılık ile ifade
edildiği bulunmuştur. Farklı alanlardaki hücreler uyaran
özelliklerinin tümü hakkında az veya çok bazı bilgiler taşırlar.
(Lance Optican & Barry Richmond ve ark.).
Renkli Görme
İnsan gözü 400-700 nm dalga boyundaki ışığı görebilir. Tek
bir dalga boyundaki ışığın karakteristik rengi vardır. Farklı
dalga boylarındaki ışıkların karışımı farklı renkler oluşturur.
Nesnenin rengi, retinal görüntüsündeki ışığın
spektral içeriğine ek olarak bağlama da
bağlıdır.
Arka planın spektral içeriğinden etkilenir.
Nesnenin rengi, yansıttığı ışığın içeriğindeki
büyük değişikliklere rağmen aynı kalabilir.
Yüzeyler üzerlerine düşen ışığı yansıtma oranları ve yansıttıkları ışığın
spektral içeriği yönünden farklıdırlar. Yansıtma fonksiyonu yüzeyin her bir
dalga boyundaki ışığı yansıtma oranını verir.
Yansıtmanın 1 olması tüm ışığı yansıttığını gösterir.
Her bir koni farklı bir fotopigment
içerir.
Fotopigmentlerin
spektral
duyarlılıkları farklıdır.
S (short) konisi kısa (420 nm), M
(medium) konisi orta (530 nm), L
(long) konisi ise hafifçe daha uzun
(560) dalga boylarına en iyi yanıt verir.
Duyarlı oldukları dalga boyları aralığı
geniştir ve çakışır.
Beynin
farklı
koni
tiplerinden
sinyalleri
karşılaştırması
renk
görmenin temelidir.
Üç primer ışığı değişik oranlarda karıştırarak farklı
renkler sentezlenebilir.
Görsel uyarana M hücreleri geçici olarak (transient), P hücreleri süreğen
(sustained) yanıt verirler.
P hücrelerinin reseptif alanlarının merkezi ve çevresi farklı konilerden girdi
alır. Reseptif alanlarının merkez ve çevresine çarpan ışığın dalga
boylarındaki farklılığa duyarlıdırlar. M hücreleri de konilerden girdi
almasına rağmen reseptif alanının merkez ve çevresine gelen koni
girdilerinin tipi farklı değildir. Reseptif alanı tüm koni tiplerinden girdi alır.
Renk bilgisi P hücre sistemi ile taşınır
M gangliyon hücreleri (%10) on-center
veya off-center’dırlar ve her ikisinde de
merkez ve çevrenin spektral duyarlılıkları
benzerdir ve geniştir.
P gangliyon hücrelerinin (%80) konilerden
gelen inputların organizasyonuna göre iki
alt tipi tanımlanmıştır: L ve M konilerinden
karşı (zıt) sinyaller alanlar (kırmızı-yeşil
karşı tip) ve L ve M konilerinden toplanmış
sinyallere karşı S konilerinden sinyal
alanlar (sarı-mavi karşı tip) (single
opponent hücreler).
Görme pigmenti melanopsinin ekspresyonu nedeniyle intrensek olarak
ışığa duyarlı bir tip gangliyon hücresi daha vardır.
LGN’dan primer görsel korteksin 4C ve 4A tabakalarına projeksiyon.
Bu tabakalardaki hücrelerin bazılarının P hücrelerine benzer reseptif
alanları vardır. Çoğu basit ve kompleks hücre akromatik uyaranlara en
iyi yanıtı verir. Az sayıda hücre renk değişikliklerine iyi yanıt verir.
4. tabakadan striat korteksin üst tabakalarına projeksiyon. V1’in 2. ve 3.
tabakalarındaki blob bölgelerinde double-opponent hücreler vardır.
Bunlar örneğin kırmızı renk merkeze uygulandığında uyarılırlar, bunun
opponent rengi yani yeşil merkeze uygulandığında ise inhibe olurlar.
Renk bilgisinin ayrı taşınıp taşınmadığı bilinmiyor. Blobların yüksek
seviyeli görsel alanlara spesifik projeksiyonlarının olması renk analizi
ile ilgili kortikal bir yolağın varlığını düşündürür.
Bloblardan V2’deki ince stripelara V4 alanına projeksiyon. V4
alanında görsel uyaranın rengine seçici birçok hücre bulunur.
Renkli görme farklı spektral duyarlılıkları olan en az iki tip
fotoreseptöre ihtiyaç duyar.
Koniler tercihen belirli dalga boylarına yanıt verirler. Dalga boyu ile
değişen elektriksel yanıtın formu değildir, bir fotonun absorbe
edilebilme olasılığıdır.
Sinir sistemi yalnız belirli bir koninin yanıtından onun duyarlı olduğu
dalga boyundaki zayıf bir ışıkla mı yoksa daha az duyarlı olduğu bir
dalga boyundaki daha kuvvetli bir ışıkla mı aydınlatıldığını
belirleyemez. Tek tip konisi olanlar renkleri algılayamaz.
İki reseptörlü yani dikromatik sistemler her bir dalga boyu için iki farklı
sinyal oluşturur. Beyin iki sinyali karşılaştırarak farklı dalga
boylarındaki ışığı ayırabilir. Bir nesne uzun dalga boyundaki ışığı
primer olarak yansıtıyor ise uzun dalga boyuna duyarlı koni sisteminin
yanıtı diğer sisteminkinden daha kuvvetli olur. Nesne kırmızı veya sarı
olarak görülür. Dalga boylarının farklı karışımları aynı iki sinyali
oluşturabilir. Böylece ışığı hafifçe farklı yansıtan yüzeyler aynı
renkteymiş gibi görülebilir.
İnsanda görme sistemi normalde yüzeylerin spektral özelliklerini ifade
etmek için üç koni sistemini kullanır.
Renk Körlüğü
Trikromat; normal insanlar üç sistem ile renkli görmeyi sağlar.
Dikromatlar; üç sistemden birini kaybetmiştir. Her yüzey yansıtma
fonksiyonu iki değerle tanımlanır.
L konileri yoksa protanopia, M konileri yoksa deuteranopia.
Erkeklerde oluşur sıklığı %1. Geni X kromozomunda.
S konileri kayıp ise tritanopia, 10 binde bir oluşur ve kadınlarla
erkekleri eşit oranda etkiler. Geni 7. kromozomdadır.
Anomalili trikromatlar; bir koni sisteminin spektral duyarlılığında
farklılaşma vardır. Normal L veya M koni pigmenti, spektral
duyarlılığı bunların arasında olan bir pigment ile yer değiştirmiştir.
Protanomali, deuteranomali. Sıklığı erkeklerde %7-8.